
When diving in southern Australian waters, there are few creatures more stunningly beautiful AND intelligent than the cephalopods. The entire family has the ability to rapidly change colours to camouflage, court and communicate, and they have the largest brains of any known marine invertebrate group. They exhibit a wide range of behaviours that indicate intelligence including learning and curiosity, and they are, above all, utterly charming.
Surprisingly cuttles (they are not fish, ergo I do not use the common term ‘cuttlefish’) are not globally distributed, the 100 or so known species are all found in the Old World regions, i.e. they are not found in the Americas. Of these, 26 species are found in Australia, 21 of which are endemic and 7 of which are found in SA waters, including Sepia apama, the giant Australian cuttle (Top picture). It has been observed from Moreton Bay, Qld (~27°S) to Ningaloo Reef, WA (~22°S) at depths of 1-100 m.
Before I launch into how glorious S. apama is, how unique their winter spawning aggregation is and how little we actually know about this animal, I will outline some basic cephalopod biology because they are incredible creatures and should be appreciated for all their amazing adaptations.
Cuttles belong to the group of cephalopods which have reduced and internalised, or lost, their shells, as opposed to the ancient Nautilus. The group is known as the coleoid cephalopods. The modified shell inside cuttles, known as cuttlebone or a ‘sepion’ and made of calcium carbonate, allows cuttles to live in the water column like a fish, able to accurately and quickly control their buoyancy by changing the quantity of gas in the tiny chambers of the sepion.
Cuttles have two modes of swimming: finning and jet propulsion. The lateral fins can be used for hovering and swimming. Jet propulsion is achieved by sucking water into the body cavity and then expelling it at high speed through a funnel underneath the body. The funnel can turn to direct the movement away from a threat coming from any direction. To further aid escape from predators, cuttles, like squid, can squirt a cloud of black ink to shield their escape and confuse the predator.
Cuttles have eight arms and two longer tentacles. The two tentacles are usually kept tucked in two pockets under the eyes, and are used to ambush prey. After sneaking up on a suitable prey item, the tentacles are shot out at high speed to seize the prey before it can escape. Cuttles consume a wide variety of animals including crustaceans. To get at the yummy flesh inside the hard exoskeleton, cuttles have a parrot-like, keratinous beak that is large and strong.
One of the most endearing and stunning features of cuttles is their ability to rapidly change colour and skin texture. Any divers reading this who have surprised, confronted or played with a cuttle will have experienced this remarkable ability and been amazed by it. To enhance their colour-changing ability cuttles have muscular ‘papillae’ in their skin that they can raise so they look wrinkly and blend in superbly with surrounding algae or rocks.
Layers of different cells – chromatophores, iridophores and leucocytes (Fig. 1), produce colour patterns. Chromatophores are muscular pigment-containing cells, which when contracted, expose a large surface area of pigment, and when relaxed show no pigment. Chromatophores come in three colour types: yellow/orange, red/orange and brown/black. They are under the direct nervous control of the brain. Lying under the chromatophores are the iridophores, mirror-like crystalline structures that reflect blue and green. Under these are the leucocytes, responsible for producing white.
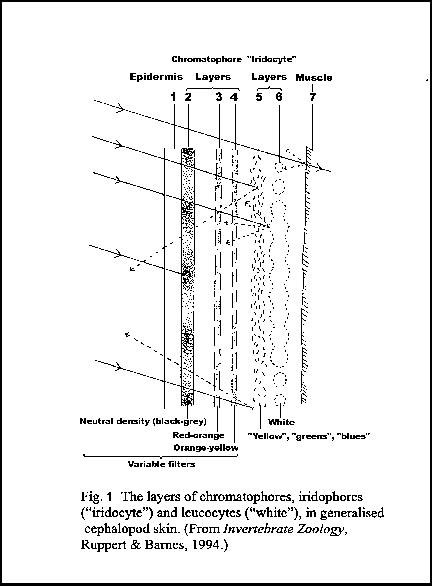
FIGURE 1
The density of chromatophores can range from 8 per mm2 in Loligo species (squid), to 50 per mm2 in Sepia species (cuttle), to 250 per mm2 in Octopus species. There is an inverse relationship between the size of chromatophores and density, therefore cephalopods like Loligo that have fewer and larger chromatophores cannot display the finer patterning seen in Sepia and Octopus.
The large and complex eyes of cephalopods correspond to large optical lobes in the brain. Despite their ability to produce an astonishing range of colour, it is widely believed by scientists that most cephalopods are colour-blind. It is proposed that instead of responding to the wavelengths of the colours in their environment, they respond to the contrast between the colours. There is evidence to support this theory. Of the 24 cephalopod species whose retinas have been examined, 23 contained only one visual pigment – rhodopsin (wavelength maximum ~480nm, depending on species). The other species, a type of squid, has 3 visual pigments, indicating that it may well see in colour. With a single visual pigment it may still be possible to distinguish colour wavelengths if the retina contains multiple banks of receptors, such as the double layers of rods found in some deep-sea fish. Light reaching the second layer is different from light entering the eye because it has already passed through the first layer of receptors and some has been absorbed. This arrangement has only been found in the same species of squid that also has the 3 visual pigments. (See Marshall & Messenger (1996) for a detailed colour-blindness experiment on a cuttle species.)
Back to Sepia apama. It ranks as one of the largest cuttle species in the world. However, we must be careful: when talking about size, many journalists are tempted to say that S. apama measure over a metre, however this measurement includes the arms and tentacles, which can stretch out to almost double the length of the animal. Correct scientific measurement uses the dorsal mantle length (ML, Fig. 2), i.e. from the front of the mantle between the eyes to the posterior tip of the mantle. The largest animals measured in SA had mantle lengths up to ~40 cm, but in New South Wales they have been measured up to ~60 cm and weigh over 5 kgs. There is no explanation for this size difference, it could be different environmental conditions, or possibly different species, but the research has not yet been done to confirm or deny either of these possibilities. The largest males are always larger than the largest females.
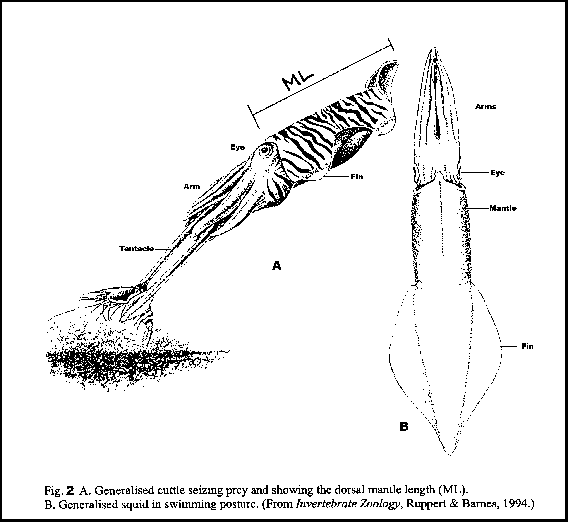
FIGURE 2
In South Australia, the giant cuttle is best known for its unique spawning aggregation that takes place every winter around an area known as Black Point, near Whyalla. Thousands of cuttles congregate over shallow inshore rocky reef. So shallow in fact that it is possible to enjoy this unique event simply by snorkeling, thus making it accessible to non-divers. Nowhere else in Australia has this type of behaviour been observed and in fact has never been recorded in any other cuttle species.
Around the end of April, beginning of May the numbers start to build up, then during August they disappear. Nobody knows where they come from, how they get there or where they go to when it’s over. SARDI scientists recently recorded densities as high as 70 cuttles per 100 m2 during the height of the season. It is not known what brings the cuttles to Whyalla each year. One suggestion is that the females need to attach their eggs to some form of hard substrate, however at this stage scientists can only guess at the reason.
In other areas of their distribution the giant cuttle has been described by recreational divers as being solitary animals, inhabiting caves and overhangs. The males appear to be very territorial and fiercely protect dens in order to attract females searching for a place to lay eggs. This kind of den-guarding behaviour has even been observed in Edithburgh. This makes the spawning aggregation at Black Point even more unique. Instead of guarding territory, males fiercely guard a chosen female. Having mated, the male aggressively fends off other males attempting to approach the female for mating, in this way he will ensure that his is the only sperm to fertilise the female’s eggs. In the meantime the female, having chosen an appropriate position for her eggs, will begin laying with great effort, apparently oblivious to the intense competition surrounding her.
There are two alternative possible explanations for this different behaviour: 1) there are very few distinct caves or overhangs for cuttles to occupy in the area, or 2) the high numbers of cuttles in one small area might prevent the majority of animals from claiming and maintaining a territory.
Competition for females is intense at Black Point with males outnumbering females 4-8:1, depending on the time during the season, and usually only the largest males could successfully court and mate. The majority of males are small sneaker or satellite males, who without a female, seek out a loosely-guarded female and attempt to mate while the ‘owner’ is distracted (this behaviour is sometimes called ‘sneaky sex’ and is seen in many other animal groups).
There are several different behaviour types that can be seen during this winter spawning time and it can often be difficult to distinguish males and females unless females are laying eggs. Even the animals themselves cannot distinguish male and female unless they display to one another. Because of the large number of males at Black Point during winter, there are many males displaying to one another to see if they are male or female and to compete with one another for a solitary, unguarded female, these are called agonistic bouts.
When males of approximately equal size approach one another, there are ‘flamboyant’ body displays, they align their bodies side by side and stretch and flare out their arms, displaying the familiar moving zebra pattern, in this way they can ‘size-up’ one another – this is the first stage of the bout. In this way they can assess if they are both male, and which is the bigger. Usually the smaller of the two will give in and move away.
If the bout continues, the next stage is called ‘flare-to-bite’. The larger male will turn to the smaller one and flare out his fourth arms as a warning to the smaller male. If the smaller male still does not back down, the larger male may move in and attempt to grab the smaller male, who will then roll out of the arms to escape.
These displays also occurred between males who already possess a female each. If approached by a very small male attempting to steal his female, a large male will merely brush the contender aside aggressively. However, the constant attention needed to guard a female from up to 6-8 other males, means that sometimes a small male may get the opportunity for sneaky sex, while the guarding male is busy displaying to another big male.
Displaying to a female is very short and subtle. The males use a small patch on the side of the body to display a very subtle version of the zebra pattern to the female, this is also used in sex recognition. It is completely up to the female whether she chooses to mate with a particular male or not. Mating takes place head to head for several minutes, while the male deposits, with his fourth arms, sacks of sperm (spermatophores) into a receptacle under the beak of the female. Some males if mating with a previously mated female, may flush out a previous male’s sperm from the female using his funnel. A female may mate with more than one male or repeatedly with the same male before laying eggs. Some females were ‘stolen’ from their male while mating or laying eggs, and were immediately mated.
Having completed mating, the female starts to search for a suitable place to hang her eggs. This appears to be a very strenuous process. The eggs are removed from the body cavity through the funnel, down the arms and stuck under a rock. They are fertilised one at a time inside the body. Anyone who has witnessed a female laying eggs will have seen the intense white appearance of her head and arms as they all scrunch up near the head, it looks like an incredible effort, the way that your knuckles go white when crunched in a fist.
Many cuttle species are thought to be ‘big bang’ spawners, i.e. they lay all their eggs in one go and then die, this is called semelparous. However, it appears that female S. apama may be able to lay more than one batch of eggs, with new eggs produced in the ovaries as each previous batch are laid. It is not known how many batches a female can lay during the season.
Eggs are laid individually within protective casings and attached to the underside of flat rocks in tight spaces. This provides some degree of protection from predators like fish, however, it appears that urchins may be eating the eggs. Several urchins have been caught ‘red-handed’ with partially consumed eggs in their mouths. Many urchins also had eggs caught on their spines. There are high densities of urchins in the area, but their predation rate and impact on cuttle eggs is unknown.
The rate of embryonic development is temperature dependent and is quicker with higher water temperature, thus those laid later in the season will mature quicker, due to increasing water temperatures in September, than those laid at the beginning of the season. Hatchlings are first observed breaking out of their cases in early September. They are approximately 1 cm (ML) but are virtually miniature adults capable of most functions including squirting ink and changing colours. Hatchlings immediately go to the substrate and hide, easily camouflaged.
After these months of fervent activity, adults begin to disappear during August. It is not known if they die after spawning, or if they return to summer feeding grounds to return to Black Point the next year. Sick-looking animals have been observed around Black Point towards the end of the spawning season. Some have been observed with the posterior tip of the sepion protruding through the mantle, causing the head and arms to droop. Once in this moribund state, the animals are incapable of withstanding rough weather and are often found washed up after storms.
There are many unanswered questions, some of them fairly simple:
- How old are the adults?
- How long do they live?
- How often do they reproduce?
- Where do the young cuttles live?
- How fast do they grow?
- Are the cuttles in SA related to those in WA or NSW?
To answer these question SARDI Aquatic Sciences and the University of Adelaide initiated a three-year research project to try to answer some of these questions. There is an urgency behind the research resulting from concerns raised over the lack of management of harvesting of cuttles at Black Point. Appropriate management protocols must be based on biological information, but despite its large size and common occurrence, S. apama has been little researched prior to now.
SARDI are undertaking a number of projects aimed at gaining some basic information: population surveys, tagging studies, reproductive studies and so on. The tagging studies will hopefully reveal whether the same animals are returning each year and how long each animal remains in the area. It may be that on animal may only stay a few days and move on, with a continuous cycle of animals throughout the season. However, the cuttles do not always approve of the blue plastic strip that has been inserted through its mantle and there are anecdotes that they have been observed removing the strips with their arms! Wouldn’t you?
Many divers and other concerned groups are lobbying to have the Black Point area made into a Marine Protected Area, or some kind of other sanctuary to prevent excessive harvesting in the future. If this species only mates and spawns once in its life then it is prone to population decline due to unmanaged fishing. Fortunately the SA Government created a closed area during the spawning season last year and again this year. The Whyalla City Council, in conjunction with the Whyalla Dive Club, are also looking into the steps required to have the spawning grounds listed as a World Heritage site.
For further information regarding the “Cuttlefish Capital” visit the website: http://www.cuttlefishcapital.com.au
I hope you can now appreciate a little more about these beguiling creatures and just how lucky we are to have this unique spectacle on our South Australian doorstep.
Acknowledgements
Many thanks to Karina Hall (of SARDI and Adelaide Uni.) for putting up with my incessant questions, for letting me photocopy everything and for reading and editing this article.
Bibliography
- Anon. (1998) Giant Cuttlefish. Australian Geographic. 51:72-87.
- Bramley, T. (2000) Divers want a Cuttlefish park. Marine and Coastal Community Network: South Australian Regional Ripples. 7(2): 2.
- Hall, K. (1999) Cuttlefish (Sepia apama). Fishery assessment report to PIRSA for the Marine Scalefish Fishery Management Committee. November 1999. South Australian Fisheries Assessment Series 99/00.
- Hall, K. (1998) The flamboyant and fascinating lifecycle of the giant cuttlefish. Southern Fisheries. Vol 6 (1): 20-25.
- Hall, K. and McGlennon, D. (1998) Cuttlefish (Sepia apama). South Australian Fisheries Assessment Series No. 98/9.
- Hanlon, R.T. and Messenger, J.B. (1996) Cephalopod Behaviour. Cambridge Press, Melbourne.
- Marshall, N.J. and Messenger, J.B. (1996) Colour-blind camouflage. Nature. 382: 408-409.
- Roper, C.F.E., Sweeney, M.J. and Nauen, C.E. (1984?) FAO Species Catalogue. FAO Fisheries Synopsis No. 125, Vol. 3 Cephalopods of the World.
- Watson-Russell, C. (199?) Cuttlefish of Sydney harbour. Australian Natural History. 20 (5): 159-163.